
Candida albicans on sieni, joka elää ja on olemassa luonnostaan ihmisen ohutsuolessa
Elimistössä elää luonnostaan pieni osuus ns. "huonoja sieniä ", symbioosissa bakteerikannan kanssa, mutta tämä tasapaino voi horjua ja siten epämiellyttävät reaktiot voivat alkaa.
Monet tekijät voivat heikentävät elimistön puolustuskykyä, lähinnä huono ruokavalio, sekä muut tekijät, kuten antibioottikuurit horjuttavat tervettä bakteeri-/hiivakantaa...
... infektio, eli kandidiaasi on Candida albicans -hiivasienen epänormaali kasvu.
... infektio, eli kandidiaasi on Candida albicans -hiivasienen epänormaali kasvu.

Candida etsii elimistöstä ravinnokseen glukoosia (rypälesokeri) ja estrogeenia, käyttäen apunaan kitiinikuorisia, nopeasti eteneviä rihmoja,
(vrt. kasvien juuristo) kuva.
Kun Candida on vallinnut pitkään elimistössä se aiheuttaa toisia ja yhä haitallisimpia sairauksia.
Kroonisessa vaiheessa esiintyy vakavia oireita, kuten sieni -infektiot emättimessä ja peniksessä, kuiva kutiseva iho, ihottumat, nivelkipu , vatsan turvotus, nukkumisongelmat, ärtyvä suoli, närästys, suutulehdus, energian puute, keskittymisvaikeudet, päänsärky, mielialan vaihtelut, kynsisieni -infektiot, ruoka-aineallergiat, j.n.e.
Joten kun Candida etenee patogeenisessä muodossa se tuottaa rakenteita, jotka muistuttavat kasvien juuria, ns. rootlets jotka tunkeutuvat suoliston seinämien läpi tehden reikiä niiden seiniin, (vuotavan suolen oireyhtymä).
Tämä mahdollistaa sienen, toksiinien, bakteerien ja suoliston sisällön vuotamisen verenkiertoon, joka puolestaan äidinmaidon kautta voi aiheuttaa vauvan suutulehduksia, ja ruoka-aineallergioita.
_
http://medicaltv.eu/
_
http://en.wikipedia.org/wiki/Fungus
(vrt. kasvien juuristo) kuva.
Candida tuottaa monia myrkyllisiä aineita, mm. gliotoksiinia, jotka aiheuttavat kipuja ja vaurioita elimistössä ja heikentävät immuunijärjestelmää.
Gliotoxin possesses immunosuppressive properties as it may suppress and cause apoptosis in certain types of cells of the immune system, including neutrophils, eosinophils, granulocytes, macrophages, and thymocytes. It also acts as an inhibitor of farnesyl transferase.
Kun Candida on vallinnut pitkään elimistössä se aiheuttaa toisia ja yhä haitallisimpia sairauksia.
Kroonisessa vaiheessa esiintyy vakavia oireita, kuten sieni -infektiot emättimessä ja peniksessä, kuiva kutiseva iho, ihottumat, nivelkipu , vatsan turvotus, nukkumisongelmat, ärtyvä suoli, närästys, suutulehdus, energian puute, keskittymisvaikeudet, päänsärky, mielialan vaihtelut, kynsisieni -infektiot, ruoka-aineallergiat, j.n.e.
Joten kun Candida etenee patogeenisessä muodossa se tuottaa rakenteita, jotka muistuttavat kasvien juuria, ns. rootlets jotka tunkeutuvat suoliston seinämien läpi tehden reikiä niiden seiniin, (vuotavan suolen oireyhtymä).
Tämä mahdollistaa sienen, toksiinien, bakteerien ja suoliston sisällön vuotamisen verenkiertoon, joka puolestaan äidinmaidon kautta voi aiheuttaa vauvan suutulehduksia, ja ruoka-aineallergioita.
_
http://medicaltv.eu/
_
http://en.wikipedia.org/wiki/Fungus
Candida albicans hyökkää immuunipuolustuksen ytimeen
(Candida albicans SAB Symptoms Of Leaky Gut Syndrome Candida)
 Hiivat ovat eukaryoottisia mikro-organismeja jotka luokitellaan kuuluviksi "sienten valtakuntaan", jossa on 1500 lajia.
Hiivat ovat eukaryoottisia mikro-organismeja jotka luokitellaan kuuluviksi "sienten valtakuntaan", jossa on 1500 lajia.Tässä kuvataan nyt (arviolta 1% kaikista sienilajeista).
Hiivat ovat yksisoluisia , vaikka jotkut hiivalajien muodot voivat tulla multisellulaarisiksi, muodostamalla kytkettyjä orastavia soluja, joita kutsutaan pesudohyphoiksi tai vääriksi rihmastoiksi.
Hiivan koko voi vaihdella suuresti lajeittain, tyypillisesti mitattuina 3-4 um in halkaisijaltaan , vaikka jotkut hiivat voi saavuttaa yli 40 mikrometriä. Useimmat hiivat lisääntyvät suvuttomasti.
Käymisessä hiiva, Saccharomyces cerevisiae, muuntaa hiilihydraatteja hiilidioksidiksi ja alkoholiksi - tuhansia vuosia hiilidioksidia on käytetty leivonnassa ja alkoholia alkoholijuomissa.
Hiiva on myös keskeisen tärkeä malliorganismi modernin solubiologian tutkimuksessa, ja on yksi eniten tutkittuja eukaryoottisia mikro-organismeja.
Tutkijat ovat käyttäneet sitä kerätäkseen tietoa eukaryoottisolun biologiasta ja lopulta ihmisen biologiasta.
Muut hiivalajit, kuten Candida albicans, ovat opportunistisia (keinoja kaihtamattomia) taudinaiheuttajia ja voivat aiheuttaa infektioita ihmisillä.
Hiivat eivät ole yksi ainoa taksonominen tai fylogeneettiseen ryhmittymä.
Termiä "hiiva" pidetään usein synonyymina Saccharomyces cerevisiae : lle.
_
http://translate.googleusercontent.c...6BVSM-8xC_ojHQ
_
http://en.wikipedia.org/wiki/Yeast
_
What are yeast?
http://wiki.yeastgenome.org/index.php/What_are_yeast%3F
____________________________________
Candidan ja hiivan rakenne
Joskus hiiva asuu ihmiskehon suolistossa (sisäympäristössä), eikä yleensä aiheuta ongelmia.
 Mutta johtuen erilaisista ympäristötekijöistä se siirtyy yksisoluisesta, tai hiiva muodosta invasiiviseen monisoluiseen muotoon, joka tunnetaan Candida: na ja alkaa lisääntyä erittäin nopeasti.
Mutta johtuen erilaisista ympäristötekijöistä se siirtyy yksisoluisesta, tai hiiva muodosta invasiiviseen monisoluiseen muotoon, joka tunnetaan Candida: na ja alkaa lisääntyä erittäin nopeasti.
Levitessään se rakentaa suojaksen helicobacter pylorin kaltaisen, biokalvon (biofilmi). Näiden biokalvojen paksuus voi vaihdella 25-450 mikrometriä ja niitä alkaa ilmetä kolonisaation ensimmäisen 24 tunnin kuluessa
Biokalvo koostuu selluloosasta, joka on kasvien rakenneosa. Polynukleotidit ovat ensisijaisesti sen DNA ja RNA materiaalia, polypeptidit, jotka ovat proteiineja, sekä polysakkarideja, jotka ovat hiilihydraatteja . Biokalvo sisältää myös fibrinogeenia, jotka ovat samoja materiaaleja, jotka elimistö käyttää veressä haavojen hyytymiseen.
 Nämä materiaalit on kiinnitetty toisiinsa lignandeilla, joilla on tahmeutta, samoja ominaisuuksia kuin vehnän lektiineillä.
Nämä materiaalit on kiinnitetty toisiinsa lignandeilla, joilla on tahmeutta, samoja ominaisuuksia kuin vehnän lektiineillä.
Eräs rakenne invasiivisen Candidan kasvaessa on pehmeä valkoinen kolonisaatio, ja toinen kasvaa sauvamaisesti harmaalla alustalla. Tämä voi olla samaa lajia Candida, ja se voi sopeutumismenetelmänään vaihtaa muotoaan.
Toisen Candida -kannan tiedetään käyvän läpi muodonvaihtoa, tuottaen seitsemää (7) erilaista kasvualustaa.
Tämä vaihtokyky on palautuva ja periytyy sukupolvelta toiselle.
Tutkimus osoitti myös, että rakenteelliset kromosomien
uudelleenjärjestelyt ovat eräs tapa geneettisen
monimuotoisuuden säilyttämiseksi ja on tämän sienen
sopeutumisstrategia.
Candida rihmasto monistuu suvullisesti, (reproduce sexually),
koska ytimellä on kaksi kopiota kromosomeista, luodakseen
kaksi erillistä solua sporangialla, kun ”mies” ja ”nainen”
kohtaavat. Jos nyt voisit kutsua sitä mieheksi ja naiseksi,
eivät todellakaan ole, ennemminkin kaksi solua jakaantuu
itsenäisesti, liittyäkseen ja luodakseen uusia soluja .
Hiiva on yksisoluinen rakenne Candida albicans: ista, ennen kuin siitä tulee monisoluisnen Candida, joita on luokiteltu yli 1000 lajia . Hiiva voi elää joko anaerobisesti tai aerobisesti.
Ilman happea ne tuottavat energiaa muuntamalla sokerit (maksan tuottaman glykoosin) hiilidioksidiksi ja etanoliksi, jos ihmettelet, miksi sinulla on aivot kuin sumussa? https://en.wikipedia.org/wiki/Warburg_hypothesis
Ne pystyvät lisääntymään suvuttomasti tai suvulliisesti ja suvuttomasti lisääntyessään solu jakautuu uutena alkuunsa (budding) ja tuottaa uusia hiivasoluja .


___________________________
See Also - katso myös:
Candida albicans infection inhibits macrophage, live cell microscopy videos

Candida albicans infection inhibits macrophage cell division and proliferationSupplementary videos 1-3
__________________________

_________________________
SYÖPÄ. Syöpäsolu ja -kasvaimet ovat poikkeuksetta väriltään valkoisia.

- oikealla tupakoitsijan keuhkosyöpä
Makrofaagit osallistuvat tuumorien kasvuun ja etenemiseen.
Ne hakeutuvat happivajauksesta (hypoksia) ja nekroosista kärsiviin kasvaimiin.
Tulehdukselliset tekijät kuten kasvain nekroosi –tekijä (TNF)-alfa jotka makrofaagi vapauttaa, aktivoi geenikytkimen, factor-kappa B.
Tämä NF-κB tunkeutuu kasvaimen nukleodiin ja käynnistää proteiinituotannon joka pysäyttää solukuoleman eli apoptoosin, edistäen samalla solun lisääntymistä ja tulehdusta.
Markofaagien on osoitettu imevän sisäänsä lukuisia kasvainsolu – tyyppejä.
Niiden lukumäärällä on vastaavuus huonoon ennusteeseen tietyissä syövissä, mukaanlukien rintasyöpä.
_
http://en.wikipedia.org/wiki/Macrophage
_
Apoptosis, ohjelmoitu solun kuolema
http://en.wikipedia.org/wiki/Apoptosis
_
Mitoosi
Mitoosi tarkoittaa tuman ja sen sisältävän perimän jakautumista kahdeksi identtiseksi kopioksi.
http://fi.wikipedia.org/wiki/Mitoosi
Makrofaagit osallistuvat tuumorien kasvuun ja etenemiseen.
Ne hakeutuvat happivajauksesta (hypoksia) ja nekroosista kärsiviin kasvaimiin.
Tulehdukselliset tekijät kuten kasvain nekroosi –tekijä (TNF)-alfa jotka makrofaagi vapauttaa, aktivoi geenikytkimen, factor-kappa B.
Tämä NF-κB tunkeutuu kasvaimen nukleodiin ja käynnistää proteiinituotannon joka pysäyttää solukuoleman eli apoptoosin, edistäen samalla solun lisääntymistä ja tulehdusta.
Markofaagien on osoitettu imevän sisäänsä lukuisia kasvainsolu – tyyppejä.
Niiden lukumäärällä on vastaavuus huonoon ennusteeseen tietyissä syövissä, mukaanlukien rintasyöpä.
_
http://en.wikipedia.org/wiki/Macrophage
_
Apoptosis, ohjelmoitu solun kuolema
http://en.wikipedia.org/wiki/Apoptosis
_
Mitoosi
Mitoosi tarkoittaa tuman ja sen sisältävän perimän jakautumista kahdeksi identtiseksi kopioksi.
http://fi.wikipedia.org/wiki/Mitoosi
____________________________
Immuniteetti, tulehdus ja syöpä
Cell. 2010 maaliskuu 19 sekä 140 (6): 883-899.
Tulehdusreaktiot ovat ratkaisevissa rooleissa kasvainten kehittymisen eri vaiheissa, mukaan lukien alkaminen, edistäminen, pahanlaatuiseksi muuntuminen, hyökkääminen, ja etäpesäkkeet.
Tulehdus vaikuttaa myös immuunipuolustukseen ja vastustaa hoitoa.
Immuunijärjestelmän solut jotka tunkeutuvat kasvaimiin harjoittavat laaja ja dynaamisia häiriöitä syöpäsolujen kanssa ja joitakin molekyylitason tapahtumia, jotka välittävät tämän vuorovaikutuksen, on paljastunut.
Tässä tarkastelussa esitetään tärkeimmät tulehdusta ohjaavien mekanismien vaikutukset ja pidättämistä kasvainten kehittymiseen ja pohditaan uusia houkuttelevia kohteita syövän hoitoon ja ehkäisyyn.
Leukosyyttisiä kasvaimia, havaittiin jo 1900 –luvulla kun Rudolf Virchow, löysi ensimmäisen merkin mahdollisesta yhteydestä tulehdus-ja syöpäsairauksien välillä.
Silti vasta viime vuosikymmenen aikana, on saatu selvää näyttöä, että tulehduksilla on kriittinen rooli kasvainten synnyssä, ja jotkut taustalla vaikuttavat molekyylitason mekanismit on selvitetty ( Karin, 2006 ).
Tulehduksen rooli tuumorigeneesissä (kasvainten synnyssä) on nyt yleisesti hyväksytty, ja on tullut ilmi, että tulehduksellinen mikroympäristö on olennainen osa kaikkia kasvaimia, joista osassa, suoraa syy-seuraussuhdetta tulehdusta ei ole vielä todistettu ( Mantovani et al., 2008 )
_
http://www.ncbi.nlm.nih.gov/pmc/articles/PMC2866629/
_
Google -suomennos
http://translate.googleusercontent.c...ZWrNpDQclp2dBQ
Valkosolut eli leukosyytit ovat verisoluja.
Ne auttavat elimistöä torjumaan tartuntatauteja osana immuunijärjestelmää. Litrassa terveen aikuisen verta on normaalisti kolmesta kahdeksaan miljardia valkosolua eli tipassa (5-10 μl) verta on noin 50 000 valkosolua. Veren lisäksi valkosoluja on pernassa, imusuonistossa ja imusolmukkeissa. Valkosoluja on eri tyyppejä ja niiden elinikä vaihtelee parista tunnista useaan vuoteen.
Valkosolut voidaan luokitella granulosyytteihin eli jyvässoluihin ja agranulosyytteihin, joita ovat monosyytit ja lymfosyytit eli imusolut.
Monosyytti erikoistuu siirtyessään verenkierrosta kudoksiin makrofagiksi, joka kuuluu syöjäsoluihin eli fagosyytteihin. Makrofagin tehtävä ontunnistaa pintansa reseptoreilla elimistölle vieras aine tai mikrobi, niellä se ja lopulta tappaa. Makrofageja on lähes kaikissa kudoksissa, mutta eniten niitä on pernassa, luuytimessä, imusolmukkeissa ja maksassa.
Makrofagien määrä lisääntyy tulehduksissa ja infektioissa, ne toimivat sekä luontaisessa että opitussa immuniteetissa.
”Valkosoluista aiheutuvia” sairauksia
Leukemia ja lymfooma ovat syöpiä, joissa valkosolut lisääntyvät hallitsemattomasti.
_
http://en.wikipedia.org/wiki/White_blood_cell
Neutrophil, (oikealla) yleisin valkosolu –laji
Monosyytti erikoistuu siirtyessään verenkierrosta kudoksiin makrofagiksi, joka kuuluu syöjäsoluihin eli fagosyytteihin. Makrofagin tehtävä ontunnistaa pintansa reseptoreilla elimistölle vieras aine tai mikrobi, niellä se ja lopulta tappaa. Makrofageja on lähes kaikissa kudoksissa, mutta eniten niitä on pernassa, luuytimessä, imusolmukkeissa ja maksassa.
Makrofagien määrä lisääntyy tulehduksissa ja infektioissa, ne toimivat sekä luontaisessa että opitussa immuniteetissa.
”Valkosoluista aiheutuvia” sairauksia
Leukemia ja lymfooma ovat syöpiä, joissa valkosolut lisääntyvät hallitsemattomasti.
_
http://en.wikipedia.org/wiki/White_blood_cell
Neutrophil, (oikealla) yleisin valkosolu –laji

Immuunijärjestelmä
Immuunijärjestelmä on elimistön puolustusjärjestelmä vieraita taudinaiheuttajia eli patogeeneja, esimerkiksi viruksia ja bakteereja, vastaan.
http://fi.wikipedia.org/wiki/Immuuni...rjestelm%C3%A4
---
Makrofaagit, eli makrofagosyytit ovat kudoksien monosyyteistä erilaistuneita soluja.
Makrofaagit löydettiin jo v. 1884. Ihmisessä makrofaagit ovat halkaisijaltaan noin 21 mikrometriä.
Monosyytit ja makrofaagit ovat molemmat phagogyyttejä. Makrofaagi toimii sekä yleisenä immuniteetin puolustajana että auttaa herättämään erityisiä puolustusmekanismeja, kuten mukautuva immuniteetti.
Makrofaageilla on yksilöllinen kyky ”syöttää” aminohappoa, arginiiniä, joko ”tappajamolekyylille” typpioksidille, tai ”korjaavalle molekyylille”, ornitiinille.
Makrofaagit myös herättävät ”tappaja” eli Th1 ja Th2 –solujen toiminnan, joiden tehtävänä phagosyyteinä, on niellä ja syödä solujätteitä sekä sairauksia aiheuttavia vierasaiheita (patogeenejä), sekä paikallaanpysyvinä että liikkuvina soluina.
Makrofaagit myös stimuloivat lymfosyyttejä ja muita immuunipuolustus soluja patogeenejä vastaan.
Makrofaagit ovat erittäin erikoistuneita fagosyyttisiä soluja jotka hyökkäävät vieraita aineita vastaan, infektoivia mikrobeja ja syöpäsoluja vastaan tuhoamalla ja syömällä. Niitä esiintyy kaikissa elävissä kudoksissa, ja niiden tarkoitus on uudistaa, (regeneroida).

A macrophage of a mouse stretching its "arms" (pseudopodia) to engulf two particles, possibly pathogens. Trypan Blue Exclusion.
__________________________________________________________
NF-κB
NF-κB (nuclear factor kappa-light-chain-enhancer of activated B cells)
is a protein complex that controls
is a protein complex that controls transcription of DNA, cytokine production and cell survival. NF-κB is found in almost all animal cell types and is involved in cellular responses to stimuli such as stress, cytokines ,free radicals, heavy metals, ultraviolet irradiation, oxidized LDL, and bacterial or viral antigens.[1][2][3][4][5] NF-κB plays a key role in regulating the immune response to infection (κ light chains are critical components of immunoglobulins). Incorrect regulation of NF-κB has been linked to cancer, inflammatory and autoimmune diseases, septic shock, viral infection, and improper immune development.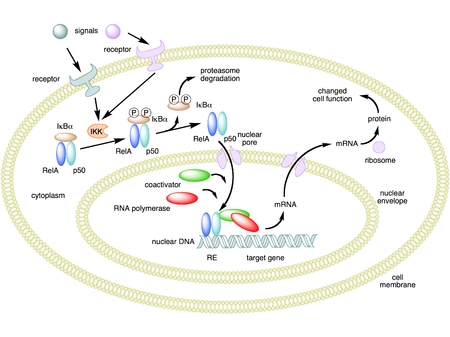
Mechanism of NF-κB action. In this figure, the NF-κB heterodimer between Rel and p50 proteins is used as an example. While in an inactivated state, NF-κB is located in the cytosol complexed with the inhibitory protein IκBα. Through the intermediacy of integral membrane receptors, a variety of extracellular signals can activate the enzyme IκB kinase (IKK). IKK, in turn, phosphorylates the IκBα protein, which results in ubiquitination, dissociation of IκBα from NF-κB, and eventual degradation of IκBα by the proteosome. The activated NF-κB is then translocated into the nucleus where it binds to specific sequences of DNA called response elements (RE). The DNA/NF-κB complex then recruits other proteins such ascoactivators and RNA polymerase, which transcribe downstream DNA into mRNA. In turn, mRNA is translated into protein, resulting in a change of cell function.[1][2][3]
Live-cell Video Microscopy of Fungal Pathogen Phagocytosis
Division of Applied Medicine, University of Aberdeen
We describe methods for live-cell video microscopy of Candida albicans phagocytosis by macrophages. These methods enable stage-specific analysis of macrophage migration, recognition, engulfment and phagosome maturation and reveal novel aspects of phagocytosis.
Video Article Chapters
6:40 - Analyzing the Live Cell Video
_
http://www.jove.com/video/50196/live...n-phagocytosis
Division of Applied Medicine, University of Aberdeen
We describe methods for live-cell video microscopy of Candida albicans phagocytosis by macrophages. These methods enable stage-specific analysis of macrophage migration, recognition, engulfment and phagosome maturation and reveal novel aspects of phagocytosis.
Video Article Chapters
6:40 - Analyzing the Live Cell Video
_
http://www.jove.com/video/50196/live...n-phagocytosis
_________
Important Notice
PubMed Central Canada website is now closed permanently
This article is no longer available through PubMedCentral Canada. You may continue to access the article at: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3430961/
All manuscripts along with all other content remain publicly searchable on PubMed Central (US) and Europe PubMed Central. Approximately 3,000 manuscripts authored by Canadian Institutes of Health Research (CIHR)-funded researchers have now also been added the National Research Council’s (NRC) Digital Repository and form the Canadian Institutes of Health Research 2009-2017 Collection.
Stage Specific Assessment of Candida albicans Phagocytosis by Macrophages Identifies Cell Wall Composition and Morphogenesis as Key Determinants
Published: March 15, 2012.
Candida albicans is a major life-threatening human fungal pathogen. Host defence against systemic Candida infection relies mainly on phagocytosis of fungal cells by cells of the innate immune system.
Candida albicans is a major life-threatening human fungal pathogen. Host defence against systemic Candida infection relies mainly on phagocytosis of fungal cells by cells of the innate immune system.
Video S1.
- Videolla makrofaagien toiminta pysähtyy, niiden nielaistua vhreiksi värjättyä Candidaa.
- Tultuaan niellyksi, Candida muuttaa olomuotoaan rihmaiseksi ja tunkeutuu ulos makrofaagista.
- Videolla makrofaagien toiminta pysähtyy, niiden nielaistua vhreiksi värjättyä Candidaa.
- Tultuaan niellyksi, Candida muuttaa olomuotoaan rihmaiseksi ja tunkeutuu ulos makrofaagista.
Shows a representative 6 hour live video microscopy of live C. albicans being ingested by macrophages.
It further illustrates hyphal growth within macrophages and macrophage killing by hyphal Candida albicans.
Video S2.
- Makrofaagi nielee useita Candida albicans -sieniä, jotka muuttavat olomuotonsa rihmaiseksi makrofaagissa.
- Makrofaagi nielee useita Candida albicans -sieniä, jotka muuttavat olomuotonsa rihmaiseksi makrofaagissa.
The macrophage ingests multiple C. albicans, which form hyphae within the macrophage. The host macrophage initiated mitosis, but instead of completely separating, the daughter cells remained fused together.
In all cases of failed cell separation, Candida albicans hyphae were observed spanning both daughter cells.

Candida albicans infection inhibits macrophage cell division and proliferation
Fungal Genet Biol. 2012 September; 49(9): 679–680.
http://pubmedcentralcanada.ca/pmcc/articles/PMC3430961/
Important Notice
PubMed Central Canada website is now closed permanently
This article is no longer available through PubMedCentral Canada. You may continue to access the article at: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3430961/
All manuscripts along with all other content remain publicly searchable on PubMed Central (US) and Europe PubMed Central. Approximately 3,000 manuscripts authored by Canadian Institutes of Health Research (CIHR)-funded researchers have now also been added the National Research Council’s (NRC) Digital Repository and form the Canadian Institutes of Health Research 2009-2017 Collection.
___________
- Published: March 15, 2012
- https://doi.org/10.1371/journal.ppat.1002578
Altered dynamics of Candida albicans phagocytosis by macrophages and PMNs when both phagocyte subsets are present.
An important first line of defense against Candida albicans infections is the killing of fungal cells by professional phagocytes of the innate immune system, such as polymorphonuclear cells (PMNs) and macrophages.
This research highlights the significance of studying pathogen-host cell interactions with a combination of phagocytes in order to gain a greater understanding of the interactions that occur between cells of the host immune system in response to fungal invasion.
Abstract
Candida albicans is a major life-threatening human fungal pathogen. Host defence against systemic Candida infection relies mainly on phagocytosis of fungal cells by cells of the innate immune system. In this study, we have employed video microscopy, coupled with sophisticated image analysis tools, to assess the contribution of distinct C. albicans cell wall components and yeast-hypha morphogenesis to specific stages of phagocytosis by macrophages. We show that macrophage migration towards C. albicans was dependent on the glycosylation status of the fungal cell wall, but not cell viability or morphogenic switching from yeast to hyphal forms. This was not a consequence of differences in maximal macrophage track velocity, but stems from a greater percentage of macrophages pursuing glycosylation deficient C. albicans during the first hour of the phagocytosis assay. The rate of engulfment ofC. albicans attached to the macrophage surface was significantly delayed for glycosylation and yeast-locked morphogenetic mutant strains, but enhanced for non-viable cells. Hyphal cells were engulfed at a slower rate than yeast cells, especially those with hyphae in excess of 20 µm, but there was no correlation between hyphal length and the rate of engulfment below this threshold. We show that spatial orientation of the hypha and whether hyphal C. albicansattached to the macrophage via the yeast or hyphal end were also important determinants of the rate of engulfment. Breaking down the overall phagocytic process into its individual components revealed novel insights into what determines the speed and effectiveness of C. albicans phagocytosis by macrophages.
Author Summary
Host defence against systemic candidiasis relies mainly on the ingestion and elimination of fungal cells by cells of the innate immune system, especially neutrophils and macrophages. Here we have used live cell video microscopy coupled with sophisticated image analysis to generate a temporal and spatial analysis in unprecedented detail of the specific effects of C. albicans viability, cell wall composition, morphogenesis and spatial orientation on two distinct stages (macrophage migration and engulfment of bound C. albicans) of the phagocytosis process. The novel methods employed here to study phagocytosis of C. albicans could be applied to study other pathogens and uptake of dying host cells. Thus, our studies have direct implications for a much broader community and provide a blueprint for future studies with other phagocytes/microorganisms that would significantly enhance our understanding of the mechanisms that govern effective phagocytosis and ultimately the innate immune response to infection.
Figures
Citation: Lewis LE, Bain JM, Lowes C, Gillespie C, Rudkin FM, Gow NAR, et al. (2012) Stage Specific Assessment of Candida albicans Phagocytosis by Macrophages Identifies Cell Wall Composition and Morphogenesis as Key Determinants. PLoS Pathog 8(3): e1002578. https://doi.org/10.1371/journal.ppat.1002578
Editor: Stuart M. Levitz, University of Massachusetts Medical School, United States of America
Received: October 14, 2011; Accepted: January 26, 2012; Published: March 15, 2012
Copyright: © 2012 Lewis et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Funding: The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript. NARG was funded by a Wellcome Trust Programme grant (080088) and an equipment grant (075470) (for DeltaVision), and by a FP7-2007–2013 grant (HEALTH-F2-2010-260338-ALLFUN). LPE is a Scottish Senior Clinical Fellow and acknowledges the support of the Chief Scientist Office (SCD/03). This work was funded by Wellcome Trust Project Grant to LPE 089930.
Competing interests: The authors have declared that no competing interests exist.
Introduction
Invasive C. albicans infection can present a serious clinical complication, especially in patients with an impaired immune system. Host defence against systemic candidiasis relies mainly on the ingestion and elimination of fungal cells by cells of the innate immune system, especially neutrophils and macrophages [1]–[3]. Despite the clinical importance of phagocytosis, this process remains poorly understood at a mechanistic level.
The fungal cell wall is the first point of contact with the innate immune system and plays an important role in recognition and phagocytosis by host immune cells [2]. It is a dynamic, highly organized organelle that determines both the shape of the fungus and its viability. The core structure of the C. albicans fungal cell wall is composed of a skeleton of polysaccharide fibrils composed of β-(1,3)-glucan that is covalently linked to β-(1,6)-glucan and chitin (a β-(1,4)-linked polymer of N-acetylglucosamine), and is designed to function as a robust exoskeleton and a scaffold for the external glycoprotein layer [4]. This outer layer consists of highly glycosylated mannoproteins that are modified by N-linked and O-linked mannosylation and phosphomannosylation [5], [6].
Another important feature of C. albicans biology thought to play a major role in host recognition is the fungus' ability to undergo reversible morphological changes between yeast, pseudohyphal, and hyphal forms in response to environmental signals [7], [8]. Its morphological plasticity is considered to be the most important virulence attribute of C. albicans [9] and plays a major role in the fungus' capacity to successfully infect many different anatomical sites of the human host. Hyphae have invasive properties that can promote tissue penetration and escape from immune cells [10], whereas yeasts are suited to dissemination in the bloodstream [9].
Phagocytic clearance of fungal pathogens may be considered to consist of four distinct stages;(i) accumulation of phagocytes at the site where fungal cells are located; (ii) recognition of fungal pathogen-associated molecular patterns (PAMPs) through pattern recognition receptors (PRRs) (reviewed in [11]; (iii) engulfment of fungal cells bound to the phagocyte cell membrane, and (iv) processing of engulfed cells within phagocytes by fusion with lysosomal vesicles to form the phagolysosome [12].
There is very limited information on how alterations in C. albicans morphogenesis or cell wall composition affect phagocyte migration towards the fungus. In contrast, a significant body of literature has identified an increasing number of PRRs and downstream signalling pathways that contribute to the recognition of fungal cells by macrophages [11], [13]. These pathways have described recognition of N-linked mannans by the mannose receptor (MR), O-linked mannans by Toll-like receptor 4 (TLR4), β-glucans by dectin-1/TLR2, and α-mannosides by galectin-3/TLR2 complexes [14]. More recently, additional PRRs have been shown to contribute to C. albicans recognition, including the scavenger receptors CD36 and SCARF1 [15], TLR9 recognition of nucleic acids [16], dectin-2 [17] and the C-type lectin mincle [18].
Comparatively little is known about the engulfment process once the fungus is tethered to the phagocyte cell membrane. However, a series of studies have shed some light on how the overall phagocytic uptake process is affected by alterations in C. albicans cell wall composition, morphogenesis and macrophage activation state [10], [19]. For example, we have recently shown that the glycosylation status of the C. albicans cell wall profoundly affected the rate of macrophage phagocytosis. Distinct patterns emerged in that phosphomannan deficient strains (mnn4Δ, pmr1Δ, and mnt3Δ mnt5Δ) were taken up at a lower rate than the wildtype or reintegrant controls, and that O-linked and N-linked mannan deficient strains are taken up at higher rates (mns1Δ and mnt1Δmnt2Δ) [10]. A study by Keppler-Ross et al. conducting competitive phagocytosis experiments suggested that macrophages displayed strong preferences for phagocytosis based on genus, species and morphology. Candida glabrata andSaccharomyces cerevisiae were taken up by J774 macrophage cells more rapidly than C. albicans, and C. albicans yeast cells were favoured over hyphal cells [20].
These studies are informative but are limited in that they assess phagocytosis in its entirety and do not break down any observed differences into individual stages of the process, such as migration, recognition or engulfment, which may be affected differentially. Furthermore, such studies assess uptake at selected time points, rather than as a continuous dynamic process, with the inherent disadvantage of ignoring temporal differences in individual stages of the phagocytosis process, which are likely to play a major role in determining the overall outcome of pathogen-host interactions in vivo.
Here we have conducted a comprehensive analysis of C. albicans phagocytosis by primary macrophages and macrophage cell lines, employing video microscopy, coupled with sophisticated image analysis tools. To assess the contribution of C. albicans cell wall glycosylation and the ability to switch from yeast to hyphal forms, we have taken advantage of a large collection of genetically and phenotypically characterized isogenic mutants of C. albicans, depleted in specific cell wall components or impaired in morphogenic switching. We show here for the first time a detailed minute by minute account of the specific effects of C. albicans viability, cell wall composition, morphogenesis and spatial orientation on two distinct stages (macrophage migration and engulfment of bound C. albicans) of the phagocytosis process. These analyses revealed that macrophage migration towards C. albicans was dependent on the glycosylation status of the fungal cell wall, but not cell viability or morphogenic switching from yeast to hyphal forms. Macrophages rapidly engulfed viable and UV-killed C. albicans, but engulfment was protracted for all glycosylation and morphogenetic mutants examined. Engulfment of hyphal C. albicans was determined by multiple components including hyphal length and spatial orientation.
Results
Macrophage migration towards C. albicans is affected by fungal cell wall glycosylation but not morphogenesis
C. albicans phagocytosis by macrophages is dependent on the C. albicans cell wall glycosylation status [10], but the question remains whether differences observed in overall uptake are a consequence of changes in migration of macrophages towards C. albicans or alterations in the engulfment process itself. Live cell video microscopy enabled examination of the individual stages of the uptake process. Representative videos are available to view in Supporting Videos S1 and S2. First we addressed the question of whether alterations in C. albicans cell wall glycosylation and morphogenesis affect migration of macrophages towards C. albicans. Primary macrophages and macrophage cell lines were challenged with glycosylation and morphogenesis defective strains of C. albicans. The strains used in this study are shown inTable 1. Briefly, the mnt1Δmnt2Δstrain is deficient in O-glycosylation [21] and has only a single O-linked mannose sugar. The mns1Δ strain has an N-glycosylation defect due to curtailed α1, 2-mannosidase activity in the endoplasmic reticulum [22] and the mnn4Δ strain has a complete loss of phosphomannan [23]. Morphogenesis defective strains included thehgc1Δ strain, a G1 cyclin mutant that is unable to form true hyphae, and efg1Δ that lacks a specific transcription factor that regulates yeast-hypha morphogenesis pathways [24], [25].
Download:
Table 1. C. albicans strains used during this study.
Migration of macrophages was assessed by live cell video microscopy using our standard phagocytosis assay 10,26, with track measurements taken at 1 min intervals over a 6 h period.Figures 1A, B and C show images derived from video microscopy depicting the track of a single macrophage migrating towards and engulfing live C. albicans (wildtype strain). Initially, the macrophage's movement appeared to be random (Figure 1A). However, dynamic analysis suggested that macrophages sensed C. albicans, accelerated and homed in on their target (Figure 1B), leading to cell-cell contact and engulfment (Figure 1C). The corresponding video is available to view in Supporting Video S3. Visual inspection of the videos suggested enhanced macrophage migration towards C. albicans glycosylation mutants, in particular themnt1Δmnt2Δstrain, compared to wildtype control.
Download:
Figure 1. Macrophage migration towards C. albicans.
Differences in macrophage velocity were observed when macrophages were cultured with different C. albicans glycosylation mutants. Tracking software was used to conduct a detailed dissection of macrophage migration dynamics. Figures 1A, B and C are snapshots from a tracking movie showing an individual macrophage (red, *) in pursuit of live wildtype C. albicans (green). Initially, the macrophage's movement appeared to be random (1A). After the macrophage had sensed C. albicans, it accelerated and homed in on its target (1B) leading to engulfment (1C). Figures 1D and E show tracking diagrams illustrating the distances travelled, directionality and velocity of J774.1 macrophages cultured with live wildtype and mnt1Δmnt2Δ glycosylation mutant, respectively. Due to the large number of macrophages tracked per video, the data was filtered to show only active macrophages with a mean track velocity greater than 1.80 µm/min. Tracks represent the movement of individual macrophages relative to their starting position, symbols indicate the location of macrophages at 1 min intervals and arrows represent directionality.
The suggestion that migration was enhanced in macrophages exposed to the mnt1Δmnt2Δ mutant strain was further supported by macrophage tracking diagrams (Figures 1D and 1E). Tracking diagrams (Figures 1D and E) illustrate the distances travelled, directionality and velocity of macrophages cultured with live wildtype and mnt1Δmnt2Δ, respectively. Due to the large number of macrophages tracked per video, the data were filtered to show only macrophages with a mean track velocity greater than that of inactive macrophages not pursuing fungal cells (1.80 µm/min). Tracks represent the movement of individual macrophages relative to their starting position, symbols indicate the location of macrophages at 1 min intervals and arrows represent directionality. These diagrams illustrate that although macrophages can migrate rapidly and for long distances when cultured with both live wildtype and themnt1Δmnt2Δ mutant, when incubated with live mnt1Δmnt2Δ a higher number of macrophages have a mean track velocity of greater than 1.80 µm/min (Figure 1E).
Quantitative analysis of average macrophage track velocity for the entire length of the observation period (6 h) showed no significant differences between wildtype (1.8 µm/min ± 0.02 SE) and yeast-locked morphogenetic mutants, but confirmed enhanced migration with UV-killed wildtype C. albicans (1.94±0.02 SE, p<0.05) and the glycosylation mutants mnt1Δmnt2Δ (2.1±0.02 SE, p<0.001), mns1Δ (2.09±0.03 SE, p<0.001) and mnn4Δ (1.96±0.03 SE, p<0.01) (Figure 2B). The macrophage average track velocity was highest for the first 30 min of the phagocytosis assay (Figure 2B). The data for this period again showed increased average track velocity for the glycosylation mutant strains mnt1Δmnt2Δ (2.68±0.04, p<0.001), mns1Δ (2.47±0.05, p<0.05) and mnn4Δ (2.52±0.07, p<0.01) when compared with wildtype (2.19±0.07) (Figure 2A). In contrast, there was no significant difference in the mean track velocity of macrophages when incubated with morphogenesis defective mutants and UV-killed wildtype. Overall track lengths were measured for the first 30 min and 6 h, and not surprisingly the data reflected the average track velocity for the wildtype and mutant strains tested (data not shown). Enhanced macrophage migration in phagocytosis assays with C. albicans glycosylation mutants was not a consequence of alterations in maximal macrophage velocity (average max velocity in µm/min: wildtype, 3.7±0.2; mnt1Δmnt2Δ, 3.6±0.1; mnn4Δ, 3.6±0.2; mns1Δ (3.1±0.2) but rather a reflection of increased macrophage activity, particularly during the first hour of the interaction assay.
Download:
Figure 2. Macrophage migration towards wildtype and mutant C. albicans.
Figures 2A and B show the mean track velocity+SE (µm/min) of J774.1 macrophages when cultured with C. albicans glycosylation and hyphal deficient mutants at 360 and 30 min time points, respectively. ns, not significant; *, p<0.05; **, p<0.01; ***, p<0.001.
Experiments with primary murine peritoneal macrophages showed a similar pattern. We observed no differences between the yeast-locked mutant strain hgc1Δ and wildtype but significantly increased average track velocity for the glycosylation mutant strain mnt1Δmnt2Δ (p<0.001) for the first 30 min and 6 h of the phagocytosis assay (Table 2). Overall the mean track velocity of peritoneal macrophages was found to be significantly lower than for J774.1 macrophages (p<0.001). However, the maximum velocity of peritoneal macrophages (3.9±0.2 µm/min) was comparable to J774.1 macrophages (3.7±0.2 µm/min). We hypothesised that the difference in mean track velocity between peritoneal and J774.1 macrophages is due in part to the peritoneal macrophages being more spread out and covering a larger surface area, therefore, reducing the need to migrate to achieve close proximity with fungal cells in the phagocytosis assay. However, experiments using the same macrophage:C. albicans ratios but lower macrophage densities confirmed that this was not the case, as mean track velocities were unchanged (Table 2). Thus, changes in C. albicans cell wall composition but not hyphal morphogenesis markedly influenced macrophage migration in vitro.
Download:
Table 2. Mean track velocity of peritoneal macrophages exposed to C. Albicans.
Macrophages rapidly engulfed viable and UV-killed C. albicans but engulfment was delayed for glycosylation and yeast-locked mutants
Effective migration of macrophages towards C. albicans is necessary to establish cell-cell contact, which is a prerequisite for initiation of the engulfment process. Next we addressed the question of whether alterations in C. albicans cell wall glycosylation and morphogenesis affected the ability and speed by which macrophages engulfed C. albicans after cell-cell contact was established. Live cell video microscopy coupled with image analysis generated a detailed minute by minute account of the engulfment process (Figures 3A, B and C). WildtypeC. albicans was shown to be rapidly engulfed by macrophages once cell-cell contact was established (Figure 3D). A three dimensional projection image confirming C. albicansphagocytosis is available in Supporting VideoS4.The average time taken for engulfment of wildtype C. albicans is 6.7±0.3 min, and the vast majority (95%) of fungal cells were engulfed within 15 min. UV-killed C. albicans yeast cells were engulfed even more swiftly, with all cells taken up within 15 min and engulfment taking an average of 4.2±0.1 min (Figure 3E).
Download:
Figure 3. Macrophage engulfment of live and UV-killed wildtype C. albicans.
Figures 3A, B, and C are snapshots taken from live cell video microscopy capturing the engulfment process. Figure 3A shows a macrophage (*) and C. albicans prior to cell-cell contact, Figure 3B shows the same cells during contact and Figure 3C shows C. albicans within the macrophage post engulfment. Scale bar, 10 µm. Figures 3D and 3Eshow the time taken for J774.1 macrophages to ingest live and UV-killed wildtype C. albicans following cell-cell contact verses the percentage of uptake events, respectively (n = 6). The majority of live C. albicans were engulfed rapidly by macrophages once cell-cell contact was established (Figure 3D). However, UV-killed C. albicans were engulfed more swiftly, with all cells internalised within 15 min (Figure 3E).
Interestingly, the rate of engulfment of all glycosylation mutant strains (Figures 4B–D) was significantly slower than that of wildtype C. albicans (mnt1Δmnt2Δ) (p<0.001); mns1Δ (p<0.01;mnn4Δ (p<0.001) (Figure 4A). The delayed engulfment was most marked for the mnt1Δmnt2Δ (13.5±0.7 min) and mnn4Δ (14.4±0.9 min) mutant strains (Figures 4B and D), as macrophages on average took twice as long to engulf these mutants than wildtype C. albicans (Figure 4A). Control strains mnt1Δmnt2Δ::MNT1 and mnn4Δ::MNN4, containing a single reintegrated copy of the corresponding deleted genes, partially restored the ability of macrophages to swiftly engulf C. albicans (data not shown). Experiments using primary thioglycollate elicited murine peritoneal macrophages and human monocyte derived macrophages also showed a significant delay for the engulfment of the glycosylation deficient mutant mnt1Δmnt2Δ and this was partially restored in the corresponding reintegrant control mnt1Δmnt2Δ::MNT1(Table 3). Engulfment of yeast-locked morphogenetic mutant strains was delayed and ultimately impaired, relative to wildtype controls in J774 macrophages (p<0.001). Firstly, the average time taken for engulfment of the hgc1Δ (Figure 5A and B) and efg1Δ (Figure 5A and C) mutant strains was significantly greater than for the wildtype control (16.2±1.4 min, 21.9±2.4 min and 6.7±0.3 min, respectively). Engulfment of approximately 1.5% of wildtype C. albicans took longer than 30 min, compared with approximately 11% and 17% for hgc1Δ and efg1Δ, respectively.
Download:
Figure 4. Macrophage engulfment of wildtype and glycosylation mutants of C. albicans.
Figures 4A, B, C and D show the time taken for J774.1 macrophages to ingest live wildtype (n = 6), mnt1Δmnt2Δ (n = 6), mns1Δ (n = 3) and mnn4Δ (n = 3) strains following initial cell-cell contact verses the percentage of uptake events. The rate of engulfment of all glycosylation mutant strains (Figures 4B–D) was significantly slower than that of wildtype C. albicans (Figure 4A).
Download:
Figure 5. Macrophage engulfment of wildtype and yeast-locked morphogenetic mutant strains of C. albicans.
Figures 5A, B and C show the time taken for J774.1 macrophages to ingest live wildtype (n = 6), hgc1Δ(n = 6) and efg1Δ (n = 3) following initial cell-cell contact plotted versus the percentage of uptake events. The average time taken for engulfment of the hgc1Δ (Figure 5B) and efg1Δ (Figure 5C) mutant strains was significantly greater than for the wildtype control (Figure 5A).
Download:
Table 3. Engulfment of wildtype and mutant of C. albicans by primary macrophages.
Secondly, we observed that approximately 2% of hgc1Δ and 41% of efg1Δ mutants that established contact with macrophages were not internalised, even after prolonged cell-cell contact. In contrast, all wildtype C. albicans yeasts were successfully engulfed following recognition. However, 66% of the efg1Δ mutant cells were eventually engulfed by neighbouring phagocytes after detachment from the macrophage they were originally in contact with. Experiments using peritoneal macrophages showed a non significant delay in the engulfment of the yeast locked mutant strain hgc1Δ (Table 3) and little evidence of detachment once cell-cell contact was established.
Thus, macrophages rapidly engulfed viable and UV-killed C. albicans after cell-cell contact was established, but engulfment was markedly slower for all glycosylation (in all macrophage subsets studied) and yeast-locked (solely in the J774 macrophage cell line) morphogenetic mutants examined.
Macrophages are more effective at engulfing yeast rather than hyphal C. albicans and engulfment is influenced by hyphal length
The data above showed that UV-killed yeast cells were engulfed more rapidly than live wildtype cells that were able to form hyphae. The accelerated engulfment of UV-killed cells raised questions about how cell morphology affects engulfment of C. albicans by macrophages. We examined the engulfment of wildtype C. albicans cells that had established cell-cell contact with macrophages in either yeast or hyphal morphology. Hyphal C. albicans cells were engulfed at a slower rate than C. albicans yeast cells (10.8±0.9 min and 5.6±0.3 min, respectively). Furthermore, the vast majority (98%) of yeast cells of C. albicans were taken up within 15 min (Figure 6A), whereas there was greater variability for hyphal cells of C. albicans, with 21% taking longer than 15 min to become engulfed (Figure 6B).
Download:
Figure 6. Role of hyphal morphogenesis in the engulfment of C. albicans by macrophages.
Figures 6A and B show the time taken for J774.1 macrophages to ingest live C. albicansin yeast and hyphal morphology plotted against the percentage of uptake events (n = 6). Hyphal C. albicans was engulfed at a significantly slower rate than the yeast form of C. albicans. Figures 6C and D show the time taken for macrophages to engulf C. albicanswith hyphae less than and greater than 20 µm, respectively (n = 6). There was no correlation between hyphal length and speed of engulfment for hyphal C. albicans of less than 20 µm length (Figure 6C). However, when hyphal length exceeded 20 µm there was significant impact on the macrophage's ability to engulf C. albicans (Figure 6D).
Next we examined whether hyphal length influenced the speed of engulfment, perhaps explaining the variations observed for hyphal C. albicans engulfment. Macrophages were capable of ingesting C. albicans with hyphae of more than twice the average diameter of macrophages (the maximum observed length of ingested hyphae was 42.9 µm), but the mean hyphal length at time of engulfment was 8.7±0.7 µm. Intriguingly, and contrary to expectations, we found no correlation between hyphal length and speed of engulfment for hyphal cells of C. albicans of less than 20 µm length (Figure 6C). However, when hyphal length exceeded 20 µm there was as significant impact on the macrophage's ability to engulf C. albicans (Figure 6D). Although macrophages engulfed C. albicans with hyphae larger than 20 µm, uptake was markedly slower with 64% of uptake events requiring more than 15 min. It is worth noting that despite having difficulty engulfing large hyphae macrophages were nonetheless persistent in their attempt to do so. Thus, macrophages were more effective at engulfing yeast cells rather than hyphal cells of C. albicans and engulfment of hyphal cells was influenced in part by hyphal length, with a cut off of 20 µm, above which macrophage engulfment was markedly impaired.
The rate of engulfment of hyphal cells of C. albicans was influenced by spatial orientation
Finally, we took advantage of the large quantity of data amassed from live cell video microscopy phagocytosis assays to address previously unanswered questions relating to how spatial orientation of C. albicans may affect the efficiency of engulfment by macrophages. First, we established that hyphal cells of C. albicans could be taken up by macrophages independent of their spatial orientation (Figure 7A). C. albicans germ tubes could be engulfed yeast-end on and germ tube apex-end on (Figures 7B and C), side-on (Figure 7D) and at an angle (Figure 7E). However, it is noteworthy that although cell-cell contact could be initiated in any orientation, the rate of engulfment was affected; C. albicans that made contact in an end-on orientation were taken up more rapidly than those engulfed at an angle or where cell-cell contact was initiated side-on (5.5±0.7 min, 8.8±0.5 min and 9.5±2.3 min, respectively). The large SE observed when C. albicans makes contact side-on can be explained by the fact that macrophages had particular difficulty ingesting large hyphae (>20 µm) in the side-on orientation. The end initially encountered by the macrophage appeared to be random. Approximately equal numbers of encounters occurred that were yeast end-on or hyphal end-on, but there was a propensity for C. albicans to be taken up more rapidly yeast-end on (8.0±0.6 min) than hyphal end-on (9.8±1.0 min).
Download:
Figure 7. The role of C. albicans spatial orientation in engulfment by J774.1 macrophages.
Figure 7A plots the time taken for engulfment of individual hyphal wildtype C. albicans in relation to the exact angles at which cell-cell contact was established (0°, side-on; 90°, end-on). C. albicans can be engulfed in any spatial orientation, including end-on, side-on and at an angle (Figure 7A). Figures 7B–E show snapshots from live cell video microscopy movies showing C. albicans being taken up in a variety of orientations, including yeast end on, (Figure 7B), germ tube apex end-on (Figure 7C), side-on (7D) and at an angle (7E). Scale bar, 10 µm; macrophage of interest, *.
Initial C. albicans orientation when establishing cell-cell contact with macrophages influenced the rate of engulfment with end-on contact of the hyphal end resulting in the most rapid engulfment. Thus, engulfment of hyphal cells of C. albicans was influenced by multiple factors including hyphal length and spatial orientation, and whether the initial encounter was by the yeast or hyphal end.
Discussion
C. albicans is a major life-threatening human fungal pathogen. Host defence against systemicCandida infection relies mainly on phagocytosis of fungal cells by cells of the innate immune system. In this study, we analysed the contribution of distinct C. albicans cell wall components and yeast-hypha morphogenesis to specific stages of phagocytosis by macrophages.
We show that macrophage migration towards C. albicans was dependent on the glycosylation status of the fungal cell wall, but not cell viability or morphogenic switching from yeast to hyphal forms. This finding was not a consequence of differences in maximal macrophage track velocity, but stems from a greater percentage of macrophages pursuing glycosylation deficientC. albicans cells during the first hour of the phagocytosis assay. The rate of engulfment of C. albicans by macrophages was significantly slower for glycosylation and morphogenesis deficient mutant strains, but enhanced for non-viable cells. Hyphal cells were engulfed at a slower rate than yeast cells, especially those with hyphae in excess of 20 µm, but there was no correlation between hyphal length and the rate of engulfment below this threshold. We show that spatial orientation of the hypha and whether hyphal C. albicans attached to the macrophage via the yeast or hyphal end were also important determinants of the rate of engulfment.
This is the first study, to our knowledge, to show that individual stages of C. albicansphagocytosis by macrophages are differentially affected by changes in C. albicans cell wall composition. Our previous work, using assays that globally assess phagocytosis, have shown increased phagocytosis of O-linked and N-linked mannan deficient strains (mns1Δ and mnt1Δmnt2Δ) [10]. Intriguingly, we show here that changes in cell wall glycosylation enhance macrophage migration towards C. albicans, but delay engulfment once cell-cell contact is established. This illustrates that standard assays do not differentiate between the individual stages of the phagocytosis process and are unable to detect significant temporal differences in migration or engulfment. For example, we show here that phosphomannan deficient cells of C. albicans were engulfed less efficiently. This effect was much less obvious in previous studies that simply evaluated phagocytosis efficiency by single time point measurements [27]. Macrophage migration was enhanced in all glycosylation mutants but most markedly in themnt1Δmnt2Δ O-glycosylation mutant. This translated into much higher overall uptake compared to the phosphomannan deficient mmn4Δ mutant and is in keeping with our previous published results [10]. It is conceivable that enhanced macrophage migration in response to the absence of O-linked (mnt1Δmnt2Δ) or N-linked mannans (mns1Δ) is a consequence of unmasking underlying β-glucans [28], [29] or electrostatic signals as a consequence of alterations in surface charge following loss of phosphomannan (mnn4Δ) [30], [31]. Observation of individual macrophage migration patterns indicated that macrophage movement was slow and random initially, but became directional towards a specific C. albicans cell, associated with a marked increase in macrophage velocity. Macrophage acceleration towards C. albicansoccurred at distances in excess of 15 µm and, therefore, suggests the presence of a chemotactic signal. Key candidates are a number of glycolipids that are known to be shed byC. albicans and are potent inducers of macrophage cytokine synthesis in vitro and in vivo [32]. We are currently conducting detailed mathematical modelling of the macrophage tracking patterns to further elucidate the hypothesis that macrophage migration towards C. albicans is affected by differences in shedding of glycolipids between wildtype and glycosylation deficient strains.
We have shown that macrophage uptake of C. albicans is a multi-step process, involving recognition and subsequent engulfment of C. albicans. C. albicans cell wall mannosylation is a key determinant in the rate of engulfment; the absence of specific PAMPs in the glycosylation mutant strains delays engulfment once cell-cell contact has been established, and this may be a consequence of differential activation of macrophage PRRs. This is in keeping with experiments in which C. albicans cell wall mutants were combined with specific macrophage receptor blocking methods that have been used to define the PAMP-PRR interactions required for cytokine induction [14], [33]. These in vitro findings are relevant to C. albicans infections in vivo, since C. albicans mutants with defects in cell wall mannosyl residues are also less virulent in experimental models of disseminated candidiasis [21], [22], [34]–[36].
Morphological plasticity is one of the hallmarks of the human fungal pathogen C. albicans [37], and its ability to switch between yeast and hyphal forms is thought to contribute to pathogenesis [24], [25], [38], [39]. C. albicans mutants that are unable to form filaments are less virulent [40], although conversely, mutants that are unable to grow as yeast are also less virulent [41]. There are conflicting reports in the literature regarding the efficiency of macrophage phagocytosis for C. albicans yeast and hyphal forms [20], [42], [43]. Here we show definitively data supporting the notion that macrophages are more effective at engulfing C. albicans yeasts. Furthermore, the use of video microscopy coupled with thorough analysis of large numbers of individual macrophage-C. albicans interactions provides a minute-by-minute account of the engulfment process, which offers detail that has not been previously available. A prime example is our observation that yeast-locked C. albicans cells were engulfed less efficiently than wildtype C. albicans. Not only was this not obvious in previous studies that simply evaluated phagocytosis efficiency by single time point measurements [10], but in addition, we observed here that delayed engulfment can result in detachment of the fungal cell and engulfment by a neighbouring macrophage. One may speculate that in vivo where phagocyte numbers are limited this may have a significant impact on pathogen clearance and that yeast locked mutantC. albicans cells have properties other than the induced phenotype that differ from wildtype yeast C. albicans cells. However, the observed delay in engulfment for yeast-locked mutant C. albicans in the macrophage cell line was almost completely abrogated in experiments using primary macrophages, underlining the importance of studying host-pathogen interactions in multiple phagocyte subsets.
The approach taken here further enabled us to dissect the complexity of engulfment of hyphalC. albicans by macrophages. We show that macrophages are capable of engulfing hyphal C. albicans in excess of 40 µm (approximately twice the diameter of macrophages) - in keeping with reports that macrophages are capable of ingesting apoptotic epithelial cells in the involuting mammary gland of similar or even larger size [44]. Hyphal length, however, does play a major role in the engulfment process of C. albicans, in that engulfment of C. albicans with hyphae in excess of 20 µm took significantly more time and phagocytosis was frequently frustrated. Interestingly, we show that below a 20 µm hyphal length threshold there was no correlation between hyphal length and the rate of engulfment. These observations are most likely related to difficulties associated with macrophages attempting to engulf very large particles. In addition to hyphal length, we have identified two other factors that influenced engulfment of hyphal C. albicans. We showed that the rate of engulfment was determined by the orientation in which C. albicans was encountered, with end-on being favourable to side-on orientation, suggesting that steric hindrance affects engulfment. We also showed that yeast end-on engulfment was more efficient than hyphal end-on encounters. This in turn may reflect differences in the wall chemistry of the hyphal tip compared to the mother cell, or be due to the efficiency of the assembly of proteins of the phagocytic cup for objects of different sizes and shapes [45].
Here we have conducted the most detailed analysis of the contribution of C. albicans viability, cell wall glycosylation and morphogenesis to phagocytosis by macrophages to date, to our knowledge. Our approach of combining live cell video microscopy with image analysis tools for the migration analysis, and minute-by-minute analysis of thousands of individual macrophage-C. albicans interactions, provides unique insight into the complexity of C. albicansphagocytosis by macrophages. The novel methods employed here to study phagocytosis of C. albicans could be applied to study other pathogens and uptake of dying host cells. Such studies would significantly enhance our understanding of the mechanisms that govern effective phagocytosis and ultimately the innate immune response to infection.
Materials and Methods
Ethics statement
All animal experiments have been conducted in strict accordance with UK Home Office guidelines. The appropriate project and personal licenses are in place PIL 60/6194 and approved by the UK Home office.
C. albicans strains and growth conditions
C. albicans serotype A strain CAI4+CIp10, hitherto referred to as the parental wildtype, was used as a control and its parent strain, CAI4, was used to construct mutants using targeted gene disruption [46]. The mutants used are listed in Table 1. C. albicans strains containing a single reintegrated copy of the corresponding deleted genes to regenerate the heterozygous genotype acted as controls. Most of the C. albicans strains used were created in house and have been described previously [21]–[25]. C. albicans strains were obtained from glycerol stocks stored at −80°C, and plated on SC-Ura plates (except hgc1Δand efg1Δ). SC-Ura plates consist of 6.9 g yeast nitrogen base without amino acids (Formedium, Norfolk, UK), 1 ml 1 M NaOH (BDH Chemicals, VWR International, Leicestershire, UK), 10 ml 1% (w/v) adenine hemisulphate salt (Sigma, Dorset, UK), 50 ml 40% D-glucose (Fisher Scientific, Leistershire, UK), 50 ml 4% SC-Ura dropout (Formedium, Norfolk, UK) and 2% (w/v) technical agar (Oxoid, Cambridge, UK) made up to 1000 ml in distilled H2O. The C. albicans morphogenetic mutantshgc1Δ and efg1Δ were grown on YPD plates consisting of 1% yeast extract (Duchefa Biochemie, Haarlem, Holland), 2% mycopeptone (Oxoid, Cambridge, UK), 2% D-glucose and 2% technical agar in distilled H2O. All plates were incubated at 30°C until colonies formed, and were then stored at 5°C.
Preparation of thioglycollate-induced peritoneal mouse macrophages
Intraperitoneal injections of 1 ml Brewer's thioglycollate broth (BD, New Jersey, USA) were administered to 8 week old female BALB/c mice. After 4 days, the peritoneal cavity of sacrificed mice was lavaged with 5 mM EDTA in 1× PBS, to harvest thioglycollate-induced macrophages. These Thio-macrophages were washed 3 times with RPMI medium 1640 (Sigma, Dorset, UK) supplemented with 10% (v/v) foetal calf serum (FCS) (Biosera, Ringmer, UK), 200 U/ml penicillin/streptomycin antibiotics (Invitrogen Ltd, Paisley, UK), 10 mM HEPES (Invitrogen Ltd, Paisley, UK) and 2 mM L-glutamine (Invitrogen, Paisley, UK). For phagocytosis assays, 1×106 thio-macrophages in 2 ml supplemented RPMI medium were seeded onto glass bottomed Iwaki dishes (VWR, Leistershire, UK) and cultured overnight at 37°C with 5% CO2. Immediately prior to experiments, RPMI medium was replaced with 2 ml pre-warmed supplemented CO2-independent medium (Gibco, Invitrogen, Paisley, UK) containing 1 µM LysoTracker Red DND-99 (Invitrogen, Paisley, UK). LysoTracker Red DND-99 is a red fluorescent dye that stains macrophage acidic organelles, enabling macrophage paths to be tracked using Volocity 5.0 software (Improvision, PerkinElmer, Coventry, UK).
Preparation of J774.1 mouse macrophage cell line and HMDM
J774.1 macrophages (ECACC, HPA, Salisbury, UK) were maintained in tissue culture flasks in DMEM medium (Lonza, Slough, UK), supplemented with 10% (v/v) FCS (Biosera, Ringmer, UK), 200 U/ml penicillin/streptomycin antibiotics (Invitrogen, Paisley, UK) and 2 mM L-glutamine (Invitrogen, Paisley, UK) at 37°C with 5% CO2. Human monocyte derived macrophages were prepared as previously described in detail [26]. For phagocytosis assays, 1×106 J774.1 macrophages in 2 ml supplemented DMEM medium were seeded onto glass based Iwaki dishes (VWR, Leistershire, UK) and cultured overnight at 37°C with 5% CO2. Immediately prior to experiments, DMEM medium was replaced with 2 ml pre-warmed supplemented CO2-independent medium (Gibco, Invitrogen, Paisley, UK) containing 1 µM LysoTracker Red DND-99 (Invitrogen, Paisley, UK).
C. albicans preparation and staining with fluorescein isothiocyanate (FITC)
Single C. albicans colonies from plates stored at 5°C were cultured in 5 ml SC-Ura/YPD medium (recipes as above, excluding technical agar) and incubated overnight at 30°C, 200 rpm. In order to determine the impact of C. albicans viability on macrophage migration and engulfment, wildtype C. albicans were killed by UV-irradiation; 100×106 fungal cells in 1 ml 1× PBS were exposed to 20 doses of UV irradiation at 100 mJ/cm2 in 6 well plates. To aid visualisation of C. albicans during phagocytosis assays, 100×106 live or UV-killed C. albicanswere stained using 1 mg/ml FITC (Sigma, Dorset, UK) in 0.05 M carbonate-bicarbonate buffer (pH 9.6) (BDH Chemicals, VWR International, Leicestershire, UK) for 10 min at room temperature in the dark. Fungal cells were washed 3 times in PBS to remove unbound FITC and resuspended in 1× PBS.
Live cell video microscopy phagocytosis assays
Our standard phagocytosis assays were performed as previously described [10]. In brief, 3×106FITC-stained C. albicans were added to 1×106 macrophages in glass based Iwaki dishes (VWR, Leistershire, UK) immediately prior to imaging. Video microscopy experiments were performed using a DeltaVision Core microscope (Applied Precision, Washington, USA) with an environmental control chamber set at 37°C. Images were captured at 1 min intervals for 6 h using an EMCCD camera. At least two independent experiments were conducted for each C. albicans strain, and at least 3 movies were analysed from each experiment. One hundred macrophages were randomly selected from each movie and their phagocytic activity determined (as below).
Analysis of live cell video microscopy movies
Volocity 5.0 imaging analysis software was used to track macrophage migration at 1 min intervals throughout the 6 h phagocytosis assay. The software enabled high throughput analysis of macrophage migration, providing detailed information on the distances travelled, directionality and velocity of thousands of individual macrophages. Data were subsequently displayed in tracking diagrams and used to calculate the mean track velocity and track length of macrophages cultured with C. albicans. These analyses enabled assessment of the affects of C. albicans viability, glycosylation status and morphology on migration.
One hundred macrophages from each movie were analysed individually at 1 min intervals throughout the 6 h phagocytosis assay.
Measurements taken include the time points at which initial cell-cell contact occurred and at which C. albicans was fully enclosed, the number of C. albicans taken up and their morphology, the orientation of hyphal C. albicans relative to the macrophage and hyphal length.
The rate of engulfment of live and UV-killed wildtype C. albicans, and glycosylation and yeast-locked morphogenetic mutant C. albicans was calculated by subtracting the time point at which initial cell-cell contact occurred from the time point at which the fungus was fully phagocytosed.
This enabled accurate assessment of the affects of C. albicans viability, glycosylation status and morphology on the speed of engulfment. C. albicans spatial orientation, morphology, hyphal length and the end of hyphal C. albicans recognised were determined to assess whether these factors impact on the rate of engulfment.
This strategy enabled in depth analysis of individual C. albicans-macrophage interactions in real time.
Measurements taken include the time points at which initial cell-cell contact occurred and at which C. albicans was fully enclosed, the number of C. albicans taken up and their morphology, the orientation of hyphal C. albicans relative to the macrophage and hyphal length.
The rate of engulfment of live and UV-killed wildtype C. albicans, and glycosylation and yeast-locked morphogenetic mutant C. albicans was calculated by subtracting the time point at which initial cell-cell contact occurred from the time point at which the fungus was fully phagocytosed.
This enabled accurate assessment of the affects of C. albicans viability, glycosylation status and morphology on the speed of engulfment. C. albicans spatial orientation, morphology, hyphal length and the end of hyphal C. albicans recognised were determined to assess whether these factors impact on the rate of engulfment.
This strategy enabled in depth analysis of individual C. albicans-macrophage interactions in real time.
Supporting Information
Video S1.
Phagocytosis of live C. albicans by J774.1 macrophages. Shows a representative 6 hour live video microscopy of live C. albicans being ingested by macrophages. It further illustrates hyphal growth within macrophages and macrophage killing by hyphal C. albicans.
(MP4)
Video S2.
Phagocytosis of live C. albicans by J774.1 macrophages. Shows a high magnification (×80) representative 6 hour live video microscopy of live C. albicans being ingested by macrophages. It further illustrates hyphal growth within macrophages and macrophage killing by hyphal C. albicans.
(MP4)
Video S3.
Macrophage tracking of C. albicans. Shows an example of a J774.1 macrophage tracking a UV-killed C.albicans yeast cell. It illustrates random macrophage movement followed by directional tracking of the fungal cell and ultimately uptake of the pathogen.
(MP4)
Video S4.
Projection movie of C. albicans phagocytosis by macrophages. Shows a 3D projection ofC. albicans ingested by macrophages. It also confirms co-localisation of green FITC stained fungal cells with lysotracker red in macrophage phagosomes.
(MOV)
Acknowledgments
We would like to thank Hector M. Mora-Montes and Gordon D. Brown for advice and useful discussions. We would also like to thank the University of Aberdeen imaging facility, in particular Kevin MacKenzie, for helpful support and advice.
Author Contributions
Conceived and designed the experiments: LEL NARG LPE. Performed the experiments: LEL JMB CG CL FMR. Analyzed the data: LEL NARG LPE. Contributed reagents/materials/analysis tools: NARG. Wrote the paper: LEL JMB NARG LPE.
References
- 1.Bistoni F, Vecchiarelli A, Cenci E, Puccetti P, Marconi P, et al. (1986) Evidence for macrophage-mediated protection against lethal Candida albicans infection. Infect Immun 51: 668–674.
- 2.Netea MG, Brown GD, Kullberg BJ, Gow NA (2008) An integrated model of the recognition of Candida albicans by the innate immune system. Nat Rev Microbiol 6: 67–78.
- 3.Sheth CC, Hall R, Lewis L, Brown AJ, Odds FC, et al. (2011) Glycosylation status of the C. albicans cell wall affects the efficiency of neutrophil phagocytosis and killing but not cytokine signaling. Med Mycol 49: 513–524.
- 4.Kapteyn JC, Hoyer LL, Hecht JE, Muller WH, Andel A, et al. (2000) The cell wall architecture of Candida albicans wild-type cells and cell wall-defective mutants. Mol Microbiol 35: 601–611.
- 5.Cutler JE (2001) N-glycosylation of yeast, with emphasis on Candida albicans. Med Mycol 39: Suppl 175–86.
- 6.Ernst JF, Prill SK (2001) O-glycosylation. Med Mycol 39: Suppl 167–74.
- 7.Moyes DL, Runglall M, Murciano C, Shen C, Nayar D, et al. (2010) A biphasic innate immune MAPK response discriminates between the yeast and hyphal forms of Candida albicans in epithelial cells. Cell Host Microbe 8: 225–235.
- 8.Cheng SC, van de Veerdonk FL, Lenardon M, Stoffels M, Plantinga T, et al. (2011) The dectin-1/inflammasome pathway is responsible for the induction of protective T-helper 17 responses that discriminate between yeasts and hyphae of Candida albicans. J Leukoc Biol 90: 357–66.
- 9.Kumamoto CA, Vinces MD (2005) Contributions of hyphae and hypha-co-regulated genes to Candida albicans virulence. Cell Microbiol 7: 1546–1554.
- 10.McKenzie CG, Koser U, Lewis LE, Bain JM, Mora-Montes HM, et al. (2010) Contribution of Candida albicans cell wall components to recognition by and escape from murine macrophages. Infect Immun 78: 1650–1658.
- 11.Brown GD (2011) Innate antifungal immunity: The key role of phagocytes. Annu Rev Immunol 29: 1–21.
- 12.Kaposzta R, Marodi L, Hollinshead M, Gordon S, da Silva RP (1999) Rapid recruitment of late endosomes and lysosomes in mouse macrophages ingesting Candida albicans. J Cell Sci 112: 3237–3248.
- 13.Netea MG, Marodi L (2010) Innate immune mechanisms for recognition and uptake ofCandida species. Trends Immunol 31: 346–353.
- 14.Netea MG, Gow NA, Munro CA, Bates S, Collins C, et al. (2006) Immune sensing ofCandida albicans requires cooperative recognition of mannans and glucans by lectin and toll-like receptors. J Clin Invest 116: 1642–1650.
- 15.Means TK, Mylonakis E, Tampakakis E, Colvin RA, Seung E, et al. (2009) Evolutionarily conserved recognition and innate immunity to fungal pathogens by the scavenger receptors SCARF1 and CD36. J Exp Med 206: 637–653.
- 16.Miyazato A, Nakamura K, Yamamoto N, Mora-Montes HM, Tanaka M, et al. (2009) Toll-like receptor 9-dependent activation of myeloid dendritic cells by deoxynucleic acids from Candida albicans. Infect Immun 77: 3056–3064.
- 17.McGreal EP, Rosas M, Brown GD, Zamze S, Wong SY, et al. (2006) The carbohydrate-recognition domain of dectin-2 is a C-type lectin with specificity for high mannose. Glycobiology 16: 422–430.
- 18.Wells CA, Salvage-Jones JA, Li X, Hitchens K, Butcher S, et al. (2008) The macrophage-inducible C-type lectin, mincle, is an essential component of the innate immune response to Candida albicans. J Immunol 180: 7404–7413.
- 19.Mora-Montes HM, Netea MG, Ferwerda G, Lenardon MD, Brown GD, et al. (2011) Recognition and blocking of innate immunity cells by Candida albicans chitin. Infect Immun 79: 1961–1970.
- 20.Keppler-Ross S, Douglas L, Konopka JB, Dean N (2010) Recognition of yeast by murine macrophages requires mannan but not glucan. Eukaryot Cell 9: 1776–1787.
- 21.Munro CA, Bates S, Buurman ET, Hughes HB, Maccallum DM, et al. (2005) Mnt1p and Mnt2p of Candida albicans are partially redundant alpha-1,2-mannosyltransferases that participate in O-linked mannosylation and are required for adhesion and virulence. J Biol Chem 280: 1051–1060.
- 22.Mora-Montes HM, Bates S, Netea MG, Diaz-Jimenez DF, Lopez-Romero E, et al. (2007) Endoplasmic reticulum alpha-glycosidases of Candida albicans are required for N glycosylation, cell wall integrity, and normal host-fungus interaction. Eukaryot Cell 6: 2184–2193.
- 23.Hobson RP, Munro CA, Bates S, MacCallum DM, Cutler JE, et al. (2004) Loss of cell wall mannosylphosphate in Candida albicans does not influence macrophage recognition. J Biol Chem 279: 39628–39635.
- 24.Zheng X, Wang Y, Wang Y (2004) Hgc1, a novel hypha-specific G1 cyclin-related protein regulates Candida albicans hyphal morphogenesis. EMBO J 23: 1845–1856.
- 25.Lo HJ, Kohler JR, DiDomenico B, Loebenberg D, Cacciapuoti A, et al. (1997) Nonfilamentous C. albicans mutants are avirulent. Cell 90: 939–949.
- 26.McPhillips KA, Erwig LP (2009) Assessment of apoptotic cell phagocytosis by macrophages. Methods Mol Biol 559: 247–256.
- 27.Hobson RP, Munro CA, Bates S, MacCallum DM, Cutler JE, et al. (2004) Loss of cell wall mannosylphosphate in Candida albicans does not influence macrophage recognition. J Biol Chem 279: 39628–39635.
- 28.Klippel N, Cui S, Groebe L, Bilitewski U (2010) Deletion of the Candida albicanshistidine kinase gene CHK1 improves recognition by phagocytes through an increased exposure of cell wall beta-1,3-glucans. Microbiology 156: 3432–3444.
- 29.Mora-Montes HM, Bates S, Netea MG, Castillo L, Brand A, et al. (2010) A multifunctional mannosyltransferase family in Candida albicans determines cell wall mannan structure and host-fungus interactions. J Biol Chem 285: 12087–12095.
- 30.Erwig LP, McPhilips KA, Wynes MW, Ivetic A, Ridley AJ, et al. (2006) Differential regulation of phagosome maturation in macrophages and dendritic cells mediated by rho GTPases and ezrin-radixin-moesin (ERM) proteins. Proc Natl Acad Sci U S A 103: 12825–12830.
- 31.McKenzie CG, Koser U, Lewis LE, Bain JM, Mora-Montes HM, et al. (2010) Contribution of Candida albicans cell wall components to recognition by and escape from murine macrophages. Infect Immun 78: 1650–1658.
- 32.Jouault T, Fradin C, Trinel PA, Bernigaud A, Poulain D (1998) Early signal transduction induced by Candida albicans in macrophages through shedding of a glycolipid. J Infect Dis 178: 792–802.
- 33.Mora-Montes HM, Bates S, Netea MG, Castillo L, Brand A, et al. (2010) A multifunctional mannosyltransferase family in Candida albicans determines cell wall mannan structure and host-fungus interactions. J Biol Chem 285: 12087–12095.
- 34.Buurman ET, Westwater C, Hube B, Brown AJ, Odds FC, et al. (1998) Molecular analysis of CaMnt1p, a mannosyl transferase important for adhesion and virulence ofCandida albicans. Proc Natl Acad Sci U S A 95: 7670–7675.
- 35.Bates S, MacCallum DM, Bertram G, Munro CA, Hughes HB, et al. (2005) Candida albicans Pmr1p, a secretory pathway P-type Ca2+/Mn2+-ATPase, is required for glycosylation and virulence. J Biol Chem 280: 23408–23415.
- 36.Bates S, Hughes HB, Munro CA, Thomas WP, MacCallum DM, et al. (2006) Outer chain N-glycans are required for cell wall integrity and virulence of Candida albicans. J Biol Chem 281: 90–98.
- 37.Lu Y, Su C, Wang A, Liu H (2011) Hyphal development in Candida albicans requires two temporally linked changes in promoter chromatin for initiation and maintenance. PLoS Biol 9: e1001105.
- 38.Gow NA, Brown AJ, Odds FC (2002) Fungal morphogenesis and host invasion. Curr Opin Microbiol 5: 366–371.
- 39.Sudbery PE (2011) Growth of Candida albicans hyphae. Nat Rev Microbiol 9: 737–748.
- 40.Kadosh D, Johnson AD (2005) Induction of the Candida albicans filamentous growth program by relief of transcriptional repression: A genome-wide analysis. Mol Biol Cell 16: 2903–2912.
- 41.Saville SP, Lazzell AL, Monteagudo C, Lopez-Ribot JL (2003) Engineered control of cell morphology in vivo reveals distinct roles for yeast and filamentous forms of Candida albicans during infection. Eukaryot Cell 2: 1053–1060.
- 42.Marcil A, Gadoury C, Ash J, Zhang J, Nantel A, et al. (2008) Analysis of PRA1 and its relationship to Candida albicans- macrophage interactions. Infect Immun 76: 4345–4358.
- 43.d'Ostiani CF, Del Sero G, Bacci A, Montagnoli C, Spreca A, et al. (2000) Dendritic cells discriminate between yeasts and hyphae of the fungus Candida albicans. Implications for initiation of T helper cell immunity in vitro and in vivo. J Exp Med 191: 1661–1674.
- 44.Monks J, Henson PM (2009) Differentiation of the mammary epithelial cell during involution: Implications for breast cancer. J Mammary Gland Biol Neoplasia 14: 159–170.
- 45.Champion JA, Mitragotri S (2006) Role of target geometry in phagocytosis. Proc Natl Acad Sci U S A 103: 4930–4934.
- 46.Murad AM, Lee PR, Broadbent ID, Barelle CJ, Brown AJ (2000) CIp10, an efficient and convenient integrating vector for Candida albicans. Yeast 16: 325–327.
http://www.ncbi.nlm.nih.gov/pubmed/24169578
Candida albicans escapes from mouse neutrophils
- Candida albicans pakenee hiiren neutrofiileiltä !
http://www.jleukbio.org/content/earl...;jlb.0213063v1
--
Estrogen-binding protein in Candida albicans
Candida albicans, the most common fungal pathogen of humans, possesses an oestrogen (estrogen) -binding protein (EBP) that binds oestrogens with high affinity and specificity. The gene that encodes the EBP (CaEBP1) has been cloned and sequenced and shown to be structurally related to the old yellow enzyme from Saccharomyces cerevisiae.
_
http://europepmc.org/abstract/MED/75...6NIqQra9aX6I.4
Infection by Candida albicans inhibits apoptosis of human monocytes and monocytic U937 cells
Journal of Leukocyte Biology Volume 60, December 1996 737
http://www.jleukbio.org/content/60/6/737.full.pdf
___
Candida albicans morphogenesis is influenced by estrogen Conversion of Candida albicans from yeast to mycelial growth is believed to be associated with the organism's virulence.
http://link.springer.com/article/10....Fs000180050094
___
Stage-specific gene expression of Candida albicans in human blood
Robert Koch-Institut, NG4, Nordufer 20, 13353 Berlin, Germany.
The pathogenic fungus Candida albicans commonly causes mucosal surface infections. In immunocompromised patients, C. albicans may penetrate into deeper tissue, enter the bloodstream and disseminate within the host causing life-threatening systemic infections.
_
http://www.ncbi.nlm.nih.gov/pubmed/12622810
_________

Candida albicans infection inhibits macrophage cell division and proliferation
This article has been cited by other articles in PMC.
Associated Data
1. Introduction
Candida albicans is a major life-threatening human fungal pathogen. Host defence against systemic infection relies mainly on phagocytosis of Candida by cells of the innate immune system (Gow et al., 2012). Our recent work has studied C. albicans-macrophage interactions using phagocytosis assays and live cell video microscopy coupled with sophisticated image analysis tools (McKenzie et al., 2010; Lewis et al., 2012). Such studies reveal dynamic aspects of the host-pathogen interaction that are not evident from studies based on fixed time point analyses. We have shown, for example, that C. albicans can rarely use non-lytic expulsion as a means to escape from macrophages, leaving both pathogen and host cell viable (Bain et al., 2012) and provided detailed information on how hyphal formation within macrophages leads to macrophage lysis and C. albicansescape (McKenzie et al., 2010). Here we show that infection of macrophages with C. albicans can result in failure of macrophages to complete mitosis. Instead of separating into two individual cells, daughter phagocytes remain together and can fuse to reform a single large macrophage. This intriguing and surprisingly common phenomenon may represent yet another virulence attribute of C. albicans.
2. Description of observation
Live cell video microscopy was used to study the dynamics of macrophage mitosis. Mitosis of J774.1 macrophages was observed frequently in the absence of C. albicans. During the 6 h observation period 30.8 ± 5.2% (mean ± s.e.) of macrophages underwent mitosis, and of these only 0.9 ± 0.9% failed to divide into two separate daughter cells. Video 1 shows an example of a J774.1 macrophage that successfully underwent mitosis. When macrophages were cultured with live C. albicans, 29.5 ± 5.7% of macrophages initiated mitosis; however, 35.9 ± 6.1% of 147 mitosis events examined resulted in failed cell division. In Video 2 the macrophage ingests multiple C. albicans, which form hyphae within the macrophage. The host macrophage initiated mitosis, but instead of completely separating, the daughter cells remained fused together. In all cases of failed cell separation, C. albicans hyphae were observed spanning both daughter cells. Macrophages did not persist in their attempt to undergo mitosis indefinitely. Instead macrophages fused back together on average 44.6 ± 3.3 min after initiation of cytokinesis. It is noteworthy that phagocytosis could still occur whilst macrophages were attempting to undergo mitosis.
Macrophages infected with the C. albicans glycosylation mutants, mnt1Δmnt2Δ, mns1Δ and mnn4Δ also underwent frequent post-mitotic fusion. For example, 26.5 ± 3.2% of 134 mitosis events examined failed to complete cell division when J774.1s were cultured with the mnt1Δmnt2Δ O-glycosylation mutant In contrast, when macrophages were cultured with UV-killed C. albicans 23.8% of macrophages underwent mitosis and of these, only 0.7 ± 0.4% of 143 attempts examined resulted in failed cell division. Mitosis was always successful when J774.1 macrophages were cultured with the hyphal morphogenetic-defective mutant efg1Δ. The majority of failed cell divisions observed during this study involved phagocytes that had taken up hyphal C. albicanscells, although it was also rarely seen in macrophages infected with clumps of C. albicans in the yeast phase morphology.
Mitotically active macrophages were often clustered in specific regions of cultures. Video 3 shows an example of four macrophages in close proximity failing to divide after initiating mitosis. One of the macrophages (B) is ruptured prior to completion of mitosis, whereas three other macrophages initiated mitosis, but the daughter cells remained fused together rather than separating into individual cells. All examples of post-mitotic fusion of daughter cells shown in Video 3 involved macrophages infected with hyphal C. albicans.
3. Discussion
This is the first report of inhibition of cell division in macrophages cultured with C. albicans to date. Inhibition of cell division has previously been reported with the human fungal pathogens Cryptococcus neoformans andCandida krusei (Luo et al., 2008; García-Rodas et al., 2011). Interestingly, infection with C. neoformans has also been shown to promote cell cycle progression as a strategy to spread infection (Luo et al., 2012). Failure to complete cell division in macrophages infected with hyphal C. albicans may occur due to difficulties in dividing large cargos, such as hyphae, between daughter cells and our observations suggest that hyphal size may be an important factor in this process. C. albicans hyphae may impede spindle formation or prevent the actin ring from contracting and pinching the macrophage into separate daughter cells during cytokinesis. Some microbial pathogens are known to interfere with cell cycle progression, including Clamydia trachomatis, which can selectively block cytokinesis (Greene and Zhong, 2003). Likewise, C. albicans may either have evolved a specific mechanism to prevent macrophage replication or interfere with this process indirectly due to the enlargement and distension of the phagosome. The observation that C. albicans hyphae spanned both daughter cells in all cases of failed cell separation implies mechanical rather than active inhibition of cytokinesis. Interestingly, non-lytic expulsion of hyphal C. albicans has been observed immediately prior to a host macrophage undergoing mitosis (Bain et al., 2012). It is possible that, in this instance, expulsion of C. albicansmay have enabled mitosis and cytokinesis to proceed normally.
Macrophages are pivotal components of the innate immune response to infection with C. albicans and mitosis of tissue-derived macrophages plays an important role in macrophage proliferation in infected tissues. Failure of cell division following mitosis induction may inhibit macrophage proliferation and the formation of new uninfected macrophages. Thus, the ability of C. albicans to impede daughter cell separation may be yet another mechanism employed by C. albicans to aid its survival. An alternative explanation is that this process benefits the host, in that macrophages may sense that they carry dividing fungal cells and inhibit division to prevent spreading of the infection. Interestingly, we observed failed cell division most frequently in clusters of macrophages exposed to and infected with multiple C. albicans hyphae. One may speculate that this may result in macrophage giant cell formation that may help suppress growth of large sized microbial cells. Failed cell division is likely to have profound consequences on the host response to infection with C. albicans and future studies are needed to establish the mechanism by which phagocytosis of fungal cells inhibits macrophage mitosis.
4. Methods
C. albicans strain CAI4-CIp10 (NGY152) was grown in SC-Ura medium at 30oC, as described previously (Lewis et al., 2012). For phagocytosis assays (McPhillips and Erwig, 2009; Mora-Montes et al., 2012), 1 × 106 macrophages in supplemented DMEM medium (Lonza, Slough, UK) were seeded onto glass based Iwaki dishes (VWR, Leistershire, UK) and cultured overnight at 37 °C with 5% CO2. During experiments, medium was replaced with supplemented CO2-independent medium containing 1 μM LysoTracker Red (Invitrogen, Paisley, UK). C. albicans were stained with 1 mg/ml FITC (Sigma, Dorset, UK) in 0.05 M carbonate–bicarbonate buffer (pH 9.6) for 10 min at 20 °C in the dark, washed three times, resuspended in PBS, and added to macrophages at a 3:1 ratio. Video microscopy was conducted at 37 °C with a DeltaVision Core microscope (Applied Precision, Washington, USA) and images were captured at 1 min intervals for 6 h using an EMCCD camera. Movies were visualised and edited using Volocity 5.0 imaging analysis software. Means and standard errors were calculated.
Acknowledgments
L.P.E. is a Scottish Senior Clinical Fellow and acknowledges the support of the Chief Scientist Office (SCD/03). This work was funded by Wellcome Trust Project Grant to L.P.E. 089930. N.A.R.G. was funded by a Wellcome Trust Programme Grant (080088) and an equipment Grant (075470) (for DeltaVision), and by a FP7-2007-2013 Grant (HEALTH-F2-2010-260338-ALLFUN).
Footnotes
Appendix ASupplementary data associated with this article can be found, in the online version, at http://dx.doi.org/10.1016/j.fgb.2012.05.007.
Appendix A. Supplementary material
Supplementary video 1:
Click here to view.(26M, mov)
Supplementary video 2:
Click here to view.(12K, jpg)
{kind=link}
Supplementary video 3:
Click here to view.(11K, jpg)
{kind=link}
References
Bain J.M., Lewis L.E., Okai B., Quinn J., Gow N.A.R., Erwig L.-P. Non-lytic expulsion/exocytosis ofCandida albicans from macrophages. Fung. Genet Biol. 2012 [PMC free article] [PubMed]
García-Rodas R., González-Camacho F., Rodríguez-Tudela J.L., Cuenca-Estrella M., Zaragoza O. The interaction between Candida krusei and murine macrophages results in multiple outcomes, including intracellular survival and escape from killing. Infect. Immun. 2011;79:2136–2144. [PMC free article] [PubMed]
Gow N.A.R., van de Veerdonk F.L., Brown A.J.P., Netea M.G. Candida albicans morphogenesis and host defence: discriminating invasion from colonization. Nat. Rev. Microbiol. 2012 [PMC free article] [PubMed]
Greene W., Zhong G. Inhibition of host cell cytokinesis by Chlamydia trachomatis infection. J. Infect.2003;47:45–51. [PubMed]
Lewis L.E., Bain J.M., Lowes C., Gillespie C., Rudkin F.M., Gow N.A.R., Erwig L.-P. Stage specific assessment of Candida albicans phagocytosis by macrophages identifies cell wall composition and morphogenesis as key determinants. PLoS Pathog. 2012;8(3):e1002578. [PMC free article] [PubMed]
Luo Y., Alvarez M., Xia L., Casadevall A. The outcome of phagocytic cell division with infectious cargo depends on single phagosome formation. PLoS One. 2008;3:e3219. [PMC free article] [PubMed]
Luo Y., Tucker S.C., Casadevall A. Fc- and complement-receptor activation stimulates cell cycle progression of macrophage cells from G1 to S. J. Immunol. 2012;174:7226–7233. [PubMed]
McKenzie C.G., Koser U., Lewis L.E., Bain J.M., Mora-Montes H.M., Barker R.N. Contribution ofCandida albicans cell wall components to recognition by and escape from murine macrophages. Infect. Immun. 2010;78:1650–1658. [PMC free article] [PubMed]
McPhillips K.A., Erwig L.P. Assessment of apoptotic cell phagocytosis by macrophages. Methods Mol. Biol.2009;559:247–256. [PubMed]
Mora-Montes H.M., McKenzie C., Bain J.M., Lewis L.E., Erwig L.P., Gow N.A.R. Interactions between macrophages and cell wall oligosaccharides of Candida albicans. Methods Mol. Biol. 2012;845:247–260.[PubMed]
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3430961/
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3430961/
______
DISCUSSION
C. albicans is an important human fungal pathogen that can cause superficial or potentially lethal systemic infections of mammalian hosts. A powerful approach in the dissection of aspects of the infection process has been to monitor the interaction between C. albicans cells and cultured or purified mammalian cells (1, 5).
Many of these studies have investigated the consequences of coculture of C. albicans cells and cells comprising elements of the mammalian immune system (15, 19, 36, 41).
This approach permits the study of the roles of different immune system cells in killing the pathogen and also allows the analysis of the effects of signaling molecules such as cytokines on specific components of the host-pathogen interaction. These studies require the measurement of the killing of C. albicans as well as the mammalian cells during the interaction; measurements of CFU (3, 43) and metabolic activity by dyes (20, 40) are typical strategies to determine pathogen killing.
Many of these studies have investigated the consequences of coculture of C. albicans cells and cells comprising elements of the mammalian immune system (15, 19, 36, 41).
This approach permits the study of the roles of different immune system cells in killing the pathogen and also allows the analysis of the effects of signaling molecules such as cytokines on specific components of the host-pathogen interaction. These studies require the measurement of the killing of C. albicans as well as the mammalian cells during the interaction; measurements of CFU (3, 43) and metabolic activity by dyes (20, 40) are typical strategies to determine pathogen killing.
The potential of this overall approach for the dissection of the host-pathogen interaction has been enhanced by the construction of C. albicans strains defective in specific molecular functions (3, 4, 23-26, 33). While these mutant strains provide important tools for the analysis of host-pathogen interactions, properties of the strains, such as changes in the ratio of yeast to hyphal forms or changes in cell wall characteristics leading to cell fragility or clumping of cells can bias assays such as CFU determinations. We have recently developed an end point dilution assay that provides a convenient measure of the effect of immune cells on Candida viability at low MOIs without the need for cell harvesting (33). In the present study we have applied this assay to a series of C. albicans mutants (cst20, cdc35, ras1, and cla4 mutants).
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC130362/
https://www.ncbi.nlm.nih.gov/pmc/articles/PMC130362/
___________________________
See Also - katso myös:
- Elevated enzyme found in most cancer patients, could GcMAF be the cure?
- How GcMAF Works - Pre-clinical trials & what we have learnt
- SUPPLEMENT GCMAF
- GcMAF immunotherapy: It's all about activating macrophages to do their work
- GcMAF macrophage activation therapy FAQ
- Goleic protein - vitamin supplement therapy
- GcMAF: The Latest Discovery in Natural Cancer Treatments
- GOLEIC the vitamin D binding protein destroying cancer at any stage and many other diseases
_________________________

| Brazil | for abscesses, bronchitis, chest problems, cough, diabetes, diarrhea, dysentery, edema, fever, intestinal colic, intestinal parasites, liver problems, neuralgia, nervousness, pain, parasites, rheumatism, spasms, worms |
| Caribbean | for chills, fever, flu, indigestion, nervousness, palpitations, rash, spasms, skin disease, and as a sedative |
| Curaçao | for childbirth, gallbladder problems, nervousness, and as a sedative and tranquilizer |
| Haiti | for digestive sluggishness, coughs, diarrhea, fever, flu, heart conditions, lactation aid, lice, nerves, parasites, pain, pellagra, sores, spasms, weakness, wounds, and as a sedative |
| Jamaica | for asthma, fevers, heart conditions, hypertension, lactation aid, nervousness, parasites, spasms, water retention, weakness, worms, and as a sedative |
Malaysia | for boils, coughs, diarrhea, dermatosis, hypertension, rheumatism, and to reduce bleeding |
| Mexico | for diarrhea, dysentery, fever, chest colds, ringworm, scurvy, and to reduce bleeding |
| Panama | for diarrhea, dyspepsia, kidney, stomach ulcers, worms |
| Peru | for diabetes, diarrhea, dysentery, fever, hypertension, indigestion, inflammation, lice, liver disorders, parasites, spasms, tumors, ulcers (internal), and as a sedative |
| Trinidad | for blood cleansing, fainting, flu, high blood pressure, insomnia, lactation aid, palpitations, ringworms |
| U.S.A. | for cancer, depression, fungal infections, hypertension, intestinal parasites, tumors |
| West Indies | for asthma, childbirth, diarrhea, hypertension, lactation aid, parasites, worms |
| Elsewhere | for arthritis, asthma, bile insufficiency, childbirth, cancer, diarrhea, dysentery, fever, heart problems, kidney problems, lactation aid, lice, liver disorders, malaria, pain, ringworm, scurvy, stomach problems, and as a sedative |
__________
More about Candida:
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
-
___
Treat candida and inflammations yourself
__________
More about Candida:
___
- https://graviolateam.blogspot.com/2014/11/kehon-ph-tasapaino-ja-ph-tasapainoteoria.html
- https://graviolateam.blogspot.com/2014/11/sairauden-syy-cause-of-disease.html
- https://graviolateam.blogspot.com/2014/10/maitohappobakteerit.html
- http://graviolateam.blogspot.com/2014/08/ruokasooda-ihmelaake-jun-5-2014-blogi.html
- https://graviolateam.blogspot.com/2016/11/is-cancer-caused-by-candida.html
- http://graviolateam.blogspot.com/2014/12/candida-albicans-sieni-kayttaytyy.html
- https://graviolateam.blogspot.com/2019/01/raymond-kirlew-cures-wifes-cancer-with.html
- https://graviolateam.blogspot.com/2018/05/9-candida-symptoms-3-steps-to-treat-them.html
- http://graviolateam.blogspot.com/2014/12/what-is-candida-yeast-diet.html
_
http://rain-tree.com/graviola.htm#.UtZow_RdVmU_
Presence of Compounds in Graviola (Annona muricata)
http://rain-tree.com/graviola-chemicals.pdf
_
Ethnomedical Information for Graviola (Annona muricata)
http://rain-tree.com/graviola-traditional-uses.pdf
http://rain-tree.com/graviola.htm#.UtZow_RdVmU_
Presence of Compounds in Graviola (Annona muricata)
http://rain-tree.com/graviola-chemicals.pdf
_
Ethnomedical Information for Graviola (Annona muricata)
http://rain-tree.com/graviola-traditional-uses.pdf
Ei kommentteja:
Lähetä kommentti
You are welcome to show your opinion here!